PRINCIPALES CONCEPTOS
Una población es un conjunto de individuos de la misma especie que viven en un lugar geográfico determinado (nicho ecológico) y que real o potencialmente son capaces de cruzarse entre sí, compartiendo un acervo común de genes. (poza de genes o “pool” génico).
La Genética de Poblaciones estudia la constitución genética de los individuos que componen las poblaciones (frecuencias génicas y genotípicas), la transmisión de los genes de una generación a la siguiente (gametos=nexos de unión entre una generación y la siguiente) y utilizando modelos matemáticos sencillos, cuando se considera 1 sólo locus y una sola fuerza actuando sobre la población, diseñados para individuos diploides con reproducción sexual.
Estas poblaciones, están sujetas a cambios evolutivos en los que subyacen cambios genéticos, los que a su vez están influenciados por factores como la selección natural y la deriva genética que actúan principalmente disminuyendo la variabilidad de las poblaciones, o migración y mutación que actúan aumentándola. El cambio genético que surja significa la perturbación del equilibrio. Con estos concepto quedaron instalados los cimientos de la genética de poblaciones, que no sería desarrollada hasta Chetverikov (1926) y Fisher, Seawall Wright y Haldane en los años 1930 - 32. Desde este momento, influiría también en la teoría de la evolución.
La demostración de este equilibrio es sencilla, como se muestra a continuación, e implica que las frecuencias génicas (la frecuencia de cada gen o alelo) permanecen constantes de generación en generación, siempre que la población cumpla las siguientes condiciones ideales:
La demostración de este equilibrio es sencilla, como se muestra a continuación, e implica que las frecuencias génicas (la frecuencia de cada gen o alelo) permanecen constantes de generación en generación, siempre que la población cumpla las siguientes condiciones ideales:
*Ser lo suficientemente amplia como para que todos los cambios que se produzcan en ella sigan las leyes de la estadística. Tampoco debe existir inmigración ni emigración.
*Los organismos componentes de esa población han de ser diploides y de reproducción al azar (panmixia).
*En esta población no hay mutaciones ni selección natural, de modo que los individuos tienen las mismas probabilidades de reproducirse, independientemente de sus genotipos.
*Los organismos componentes de esa población han de ser diploides y de reproducción al azar (panmixia).
*En esta población no hay mutaciones ni selección natural, de modo que los individuos tienen las mismas probabilidades de reproducirse, independientemente de sus genotipos.
Mutación: un cambio ocurrido en el genoma de una célula, que se transmite a su descendencia dando lugar a células hijas o a individuos que se denominan mutantes.La mutación es la fuente última de variación genética. Es aleatoria (independiente, no dirigida) de la función del gen. La mutación es un proceso que cambia la estructura genética de las poblaciones a un ritmo muy lento. Las tasas de mutación son muy bajas y por ello no pueden producir cambios de frecuencias (por generación) rápidos en las poblaciones.
Migración: es el movimiento de individuos entre poblaciones. Si las poblaciones difieren en frecuencias alélicas o génicas, la migración puede producir cambios importantes en las frecuencias alélicas. El movimiento de genes de una población a otra se denomina “flujo genético”. En la migración los cambios en las frecuencias alélicas son proporcionales a las diferencias de frecuencias entre la población donadora y receptora y también son proporcionales a la tasa de migración.
Dériva genética (aleatoria): Puesto que las poblaciones naturales tienen un tamaño finito, en cada generación hay un sorteo de genes durante la transmisión de gametos de los padres a los hijos que hace que las frecuencias de los alelos fluctúen de generación en generación. La deriva genética es el efecto acumulativo de esta fluctuación genética durante muchas generaciones. Si “p” ó “q” = 1, entonces ya no es posible un cambio de frecuencias porque sólo hay una variante alélica. El efecto último de la deriva genética es la fijación de uno de los alelos en la población. La tasa de fijación es inversamente proporcional al tamaño de la población (la tasa de fijación de alelos es mayor en poblaciones pequeñas).
Migración: es el movimiento de individuos entre poblaciones. Si las poblaciones difieren en frecuencias alélicas o génicas, la migración puede producir cambios importantes en las frecuencias alélicas. El movimiento de genes de una población a otra se denomina “flujo genético”. En la migración los cambios en las frecuencias alélicas son proporcionales a las diferencias de frecuencias entre la población donadora y receptora y también son proporcionales a la tasa de migración.
Dériva genética (aleatoria): Puesto que las poblaciones naturales tienen un tamaño finito, en cada generación hay un sorteo de genes durante la transmisión de gametos de los padres a los hijos que hace que las frecuencias de los alelos fluctúen de generación en generación. La deriva genética es el efecto acumulativo de esta fluctuación genética durante muchas generaciones. Si “p” ó “q” = 1, entonces ya no es posible un cambio de frecuencias porque sólo hay una variante alélica. El efecto último de la deriva genética es la fijación de uno de los alelos en la población. La tasa de fijación es inversamente proporcional al tamaño de la población (la tasa de fijación de alelos es mayor en poblaciones pequeñas).
PRINCIPIO DE HARDY WEINBERG
En el año 1908 se formuló un descubrimiento importante, por partida doble e independientemente: el matemático Hardy en Gran Bretaña y el antropólogo Weinberg en Alemania demostraron que la composición genética de una población permanece en equilibrio mientras no actúen ni la selección ni ningún otro factor y no se produzca mutación alguna. A pesar de la mezcla de genes que supone la reproducción sexual, la persistente reorganización de estos en este tipo de reproducción no cambia la frecuencia de estos en las sucesivas generaciones. Es decir, la herencia mendeliana, por sí misma, no engendra cambio evolutivo, no es un mecanismo de alteración de las frecuencias de los genes en las poblaciones.Este principio es conocido como equilibrio Hardy-Weinberg.
Dicha ley dice; “En una población panmíctica, suficientemente grande y no sometida a migración, mutación, deriva génica o selección, las frecuencias génicas y genotípicas se mantienen constantes de generación en generación”. Cuando se cumplen estas condiciones tal población se dice que está en equilibrio Hardy-Weinberg.
HETEROSIS Y HOMOCIGOSIS
Cuando cruzamos dos líneas puras ( homocigóticas) generalmente se obtiene una descendencia que es superior en casi todas las carácterísticas morfológicas y fisiológicas, así como de adaptabilidad (fitness) a cualquiera de los padres intervinientes en su formación.
Este fenómeno, descrito como heterosis por Shull (1914), estaría dado por la reaparición de la heterocigosidad en los genes que estaban al estado homocigótico en las líneas puras. Heterosis es un término utilizado en genética y en crianza y mejoramiento selectivo. También es conocido como vigor híbrido, describe la mayor fortaleza de diferentes características en los mestizos; la posibilidad de obtener "mejores" individuos por la combinación de virtudes de sus padres.
La heterosis es lo opuesto al proceso de endogamia, donde se exalta la homocigosis. Aunque se cree que la heterosis es la acción de muchos genes de pequeño efecto, la depresión homocigótica es por acción de pocos genes de gran efecto.
El término ofrece controversia, particularmente en el mejoramiento selectivo del animal doméstico, porque se prejuzga de que todos los mestizajes de plantas o animales son mejores que sus padres; y no es necesariamente verdad. Cuando un híbrido es superior a sus progenitores se habla de "vigor híbrido".
Puede pasar que el mestizo herede tales diferencias taras de sus padres que lo hagan directamente inviable para nacer. Esta es una posibilidad extrema de la "carga alélica"
La heterosis puede clasificarse en heterosis de ambos padres, donde el híbrido muestra dimensiones incrementadas del promedio paterno, y heterosis del mejor padre, el incremento dimensional es mayor al mejor de los padres. La primera heterosis es más común en la naturaleza, y más sencillo de explicar (mecanismo de dominancia génica). La heterosis son las ventajas de los animales cruzados sobre los de raza pura.
Los individuos diploides poseen en sus células dos juegos de cromosomas homólogos, uno aportado por el gameto masculino y el otro por el gameto femenino. Dado que los genes residen en los cromosomas, resulta evidente que para cada carácter el individuo tendrá dos genes. Si en ambos cromosomas homólogos reside el mismo alelo diremos que el individuo es homocigótico para ese carácter. Por ejemplo, un guisante que tenga como genes para el color AA, es homocigótico, también lo es el que tenga aa. Por el contrario, si en cada homólogo hay un alelo distinto, el individuo será heterocigótico para ese carácter. Por ejemplo, los guisantes Aa serían heterocigóticos.
La homocigosis es la condición en la cual existen dos alelos idénticos en un mismo locus de cromosomas homólogos.
Estos términos hacen referencia a lo que ocurre en individuos heterocigotos, que tienen 2 alelos diferentes en un mismo locus. Un alelo dominante anula completamente a un alelo recesivo, por lo que sólo se puede ver lo que indique el alelo dominante. Esto indica que un individuo que un alelo dominante y otro recesivo tendrá la misma característica concreta que uno con 2 copias del alelo dominante. Los alelos dominantes se designan mediante letras mayúsculas, mientras que los recesivos lo hacen con letras minúsculas.
CONSANGUINIDAD
Es la relación de sangre entre personas que descienden, sea legítimamente o ilegítimamente, de un ancestro común.
Es la relación de sangre entre dos personas: los parientes consanguíneos son aquellos que comparten sangre por tener algún pariente común, los parientes no consanguíneos son aquellos que no presentan un vínculo de sangre, pero que son parientes por un vínculo legal (matrimonio). A esta otra relación de parentesco se le denomina afinidad.
La consanguinidad tiene grados en función del número de generaciones interpuestas en el árbol genealógico. Así, la relación padre-hijo es de primer grado, mientras que la de abuelo-nieto es de segundo grado.
También se diferencia entre:
- Línea directa: se llama así a la constituida por la serie de grados entre personas que descienden una de otra.
- ascendente (progenitores, abuelos, etc.).
- descendente (hijos, nietos, etc.).
- Línea colateral: es la constituida por la serie de grados entre personas que no descienden unas de otras, pero que proceden de un tronco común (hermanos, tíos, primos, etc.).
Los parientes de primer grado, como los padres e hijos y los hermanos carnales, tienen la mitad de sus genes comunes. Luego a mayor grado de parentesco en la pareja, mayor probabilidad de que alguno de los genes defectuosos que portamos sea transmitido por ambos padres a los hijos. De acuerdo con estudios poblacionales, se sabe que el riesgo de malformaciones y muertes infantiles entre los parientes de primer grado está aumentado en un 30% con respecto a la población general no consanguínea. Entre primos hermanos este incremento es del 3%.

BIBLIOGRAFÍA
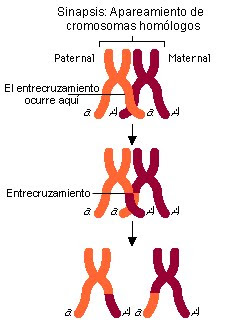
 El entrecruzamiento fue descrito por Thomas Hunt Morgán, es un proceso por el cual dos cromosomas se aparean e intercambian secciones de su ADN
El entrecruzamiento fue descrito por Thomas Hunt Morgán, es un proceso por el cual dos cromosomas se aparean e intercambian secciones de su ADN